


































Gestalt und Buntheit von Pflanzen und das ästhetische Empfinden
Wenn es um das Schöne an sich geht, verbinden wir das vielfach mit dem "Goldenen Schnitt", und dieser wiederum findet ihren Niederschlag in der antiken Architektur und Kunst. Sind die harmonischen Proportionsverhältnisse im Bau von antiken Tempeln vielleicht rein intuitiver "Natur", einem im Unterbewusstsein des Säugetiers Homo sapiens verankerten allgemein gültigen Prinzips gehorchend? Vermessungen lassen immer wieder "Goldene Schnitte" nachweisen. Eine erste, wenn auch etwas schwierig zu verstehende, Definition des "Goldenen Schnitts" geht auf Euklid (ca. 300 v.Chr.) zurück, der von einem "mittleren und äußeren Maßverhältnis" spricht. Erst im Mittelalter und in der Renaissance finden wir die Begriffe "sectio aurea" (= goldener Schnitt) und "sectio divina" (= göttlicher Schnitt). Eine mathematische Definition des "Goldenen Schnitts" ist die Teilung einer geraden Linie in der Weise, dass das Verhältnis der gesamten (ungeteilten) Strecke zum größeren Teil der (geteilten) Strecke dem Verhältnis des größeren zum kleineren Teil gleich ist (Abb. 1). Das als Zahl berechnete Teilungsverhältnis des "Goldenen Schnitts" ist eine irrationale Zahl (lässt sich nicht als Bruch ganzer Zahlen darstellen). Das mathematische Symbol ist das Φ (Phi) = 1,618033….. Die Mathematik kann uns in ein vergnügliches Erstaunen versetzen, wenn wir in punkto "Goldener Schnitt" unsere zu teilende Linie zu einem Kreis schließen (Abb. 2). Mit dem Eintrag zweier Radien an den Grenzpunkten lassen sich die entsprechenden Winkel mit einem Winkelmesser (in etwa) ermitteln. Die beiden Winkelgrade entsprechen natürlich ebenfalls irrationalen Zahlen.
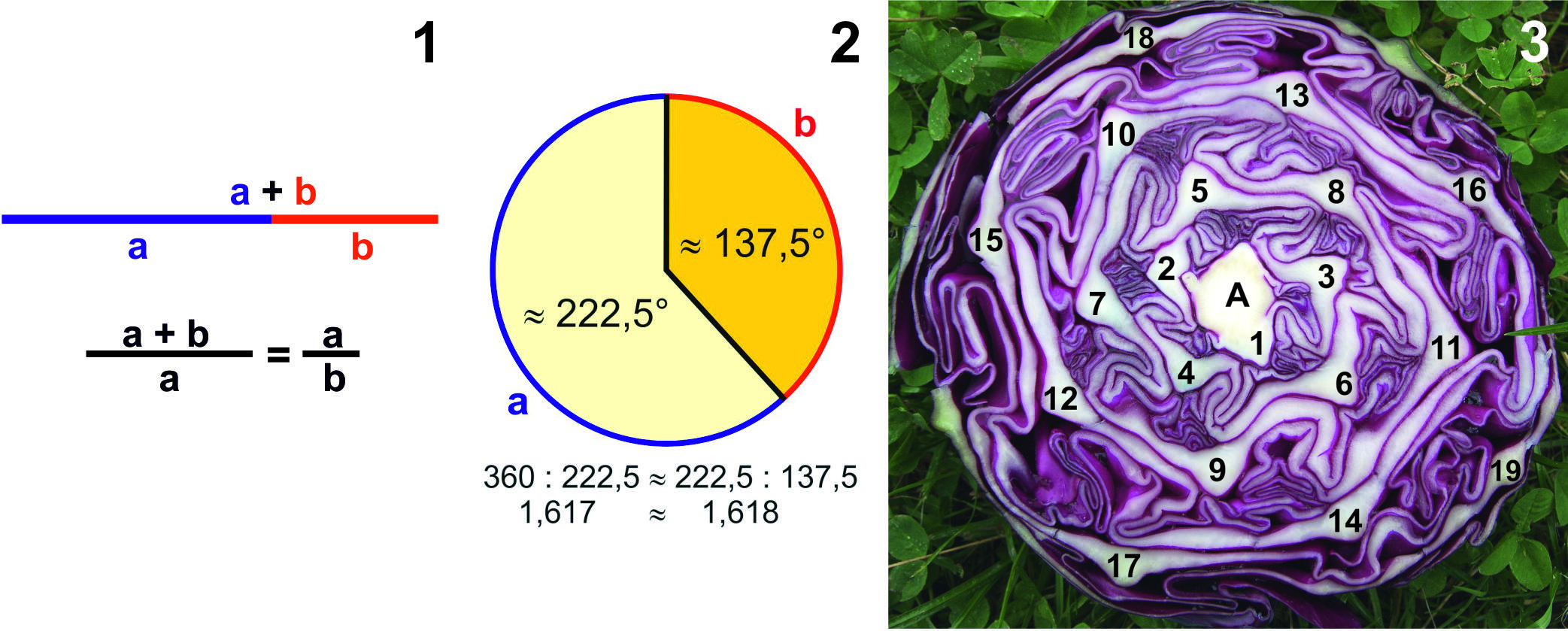
Erstaunlich ist nun, dass bei Blütenpflanzen bisweilen die raum-zeitliche Folge von Blättern oder Blütenteilen oder Blüten dem kleineren "Goldenen-Schnitt-Winkel" von ungefähr 137,5° gleichkommt und z.B. in einer Blattrosette eine gewisse Schönheit "vermittelt". Es kommen jedoch sehr oft auch bestimmte! andere Winkel vor, deren Berechnung ohne Benutzung eines Winkelmessers wir zwei genialen Köpfen des 19. Jahrhunderts, nämlich Karl Schimper und Alexander Braun, zu verdanken haben. Durch eine Bruchreihe zweier versetzter Fibonacci-Zahlenreihen (die aufeinanderfolgenden Zahlen ergeben sich bei Fibonacci durch Addition der jeweiligen beiden vorausgegangenen, nämlich 1/2 1/3 2/5 3/8 5/13 8/21 ….) erhalten wir jeweils mit 360 multipliziert die Winkelgrade. Wiederum erstaunlich ist es, dass fast ausschließlich nur die Winkel (Divergenzen) der Schimper-Braun'schen Reihe bei "Wechselständigkeit" vorkommen. Da die Differenzen der Winkelgrade rasch in der Reihe zunehmend kleiner werden, ist schon bald eine Annäherung an den Grenzwinkel erreicht. Beim Querschnitt eines Rotkohls (Abb. 3; A = Achse) sind wir bei einer 5/13-Divergenz dem Grenzwinkel schon ziemlich nahe (5/13 x 360 = 138,46…). Auf dem Kohlquerschnitt nummerieren wir von außen nach innen oder besser! von innen nach außen die Mittelrippen der in der riesigen Blattknospe gefalteten Blätter; wir drehen uns spiralförmig mit 5 Umrundungen, bis das 14. Blatt wieder mit einer neuen Spiralfolge beginnt; Nr. 14 steht genau über Nr. 1; bei der Ermittlung des Bruches darf es nicht mitgezählt werden. Zählerzahl 5 (Umdrehungszahl) ergibt sich automatisch aus der Nennerzahl 13, legt man die Schimper-Braun'sche Reihe zugrunde. Einen gewissen ästhetischen Eindruck vermitteln bei spiraliger (bzw. schraubiger) Anordnung sog. Schrägzeilen (siehe etwa 1,4,7,10,13,16,19 oder 2,5,8,11,14,17 oder 3,6,9….). Der Eindruck mag sich bei einer noch größeren Annäherung an die Limitdivergenz (kein Organ steht mehr über einem ersten) noch deutlich verstärken, so beim Romanesco, einer Variante des Blumenkohls, bei der die Seitenteile (Blütenstandsachsen) sich wiederholt einer entsprechenden Aufgliederung hingeben.
Die Regulation der Entwicklungsmuster erfolgt über komplizierte sich auf- und abbauende Hemmgradienten im Zusammenspiel mit bestimmten Größenverhältnissen in den Bildungsgeweben der Achsen zu den entstehenden Organanlagen. Alles ist optimal dicht gepackt und gehorcht dem Ökonomischen Prinzip, das möglicherweise mit dem Schönheitsprinzip identisch ist.
Ist es nur die Mathematik, die unser Schönheitsempfinden bei den Blumen bestimmt? Sind es nicht eher bizarre Formen und Düfte und vor allem die Buntheit von Blumen, die uns diese als schön empfinden lassen? Evolutionär gesehen, steht die Buntheit der Blumen in einem profanen Zusammenhang. Die Färbung der Blüten spielt nämlich als optisches Signal bei der Anlockung von bestäubenden Tieren eine außerordentlich wichtige Rolle. Die Zwitterblütenbildung, die Zusammenfassung der Pollenkörner produzierenden und Samenanlagen tragenden, gemeinsam mit dem Einschluss der Samenanlagen bei den Blütenpflanzen (Angiospermae), fand, wie Fossilfunde zeigen, wohl in der Unteren Kreidezeit, also vor 140-150 Millionen Jahren, statt. Das parasitäre Verhältnis bei den Vorläufern, den windbestäubten Nacktsamern (Gymnospermae), bei denen Insekten Pollenkörner an den männlichen Zapfenständen fraßen, wandelte sich in ein mutualistisches um, bei dem nun bei einem Blütenbesuch Tiere die Bestäubung (Übertragung der Pollenkörner auf die Narben der Fruchtblätter) besorgen können, wobei der Transport der Pollenkörner auf der Oberfläche der Insekten von diesen natürlich völlig unbeabsichtigt ist. Insekten als neues Pollen-Transportmittel werden jetzt regelrecht zur Speise eingeladen. Die Staubgefäße mit ihren Pollenkörnern waren das phylogenetisch ursprüngliche Nahrungsangebot der Blüten an die bestäubenden Insekten und wurden im Kontrast zur farbigen Blütenhülle präsentiert (Abb. 4).

Im Laufe der Evolution reduzierte sich die Zahl der Staubgefäße und damit auch die Menge an Pollenkörnern. Darüber hinaus wurde die "Belohnung" mit wertvollen proteinreichen Pollenkörnern wohl schon früh durch den vergleichsweise "billigen" zuckerreichen Nektar ergänzt oder ersetzt. Das ursprüngliche optische Signal der realen Lockspeise ging schließlich ganz verloren, als im weiteren Verlauf die Staubgefäße und der Nektar zu deren Schutz in Röhren verborgen wurden. Je tiefer Pollen und Nektar verborgen wurden, desto länger wurden – in gegenseitiger Anpassung – die Mundwerkzeuge der Insekten, um Nahrung in den Röhren zu erreichen. Damit Bestäuber trotz dieser evolutiven, vom Zufall geprägten Veränderungen auch weiterhin angelockt werden können, musste der Verlust der ursprünglichen optischen Signale, die für die Insekten weiterhin wirksam blieben, kompensiert werden. Dies erfolgte mit "Blütenmalen", optisch auffallenden Regionen einer Blüte, die als Punkt-, Flächen- oder Strichmale, aber auch als dreidimensionale Strukturen auftreten können (Abb. 5–7). Quasi werden "Wirtshausschilder" für die Insekten ins Schaufenster gestellt. Optische und olfaktorische Signale wurden zu unverwechselbaren Werbemitteln der Blüten. Zu Insekten wie Käfern, Fliegen, Bienen, Wespen, Tag- und Nachtschmetterlingen gesellten sich in der späteren Evolution Vögel und Fledermäuse hinzu. Bevorzugte Farben und Düfte sowie Unterschiede in den visuellen Fähigkeiten und im Bau der Mundwerkzeuge bzw. Zungen gaben Anlass, die Blumentypen mit eigenen sog. Syndromen ihrer entsprechenden Angepasstheiten zu charakterisieren.
Die Signale der Blumen, ihre Sprache, wird seit rund 140 Millionen Jahren von tierischen Bestäubern verstanden. Und diese erfolgreiche Partnerschaft, die für die Bestäubung von mehr als 80% der Blütenpflanzen-Arten unerlässlich ist, wird nun vom Menschen bedroht. "Pflanzenschutzmittel", speziell die Neonicotinoide, wirken nachgewiesenermaßen bei Insekten auf verhaltenssteuernde Gehirnprozesse wie Wahrnehmen, Lernen, Erinnern, Orientieren, Navigieren, Kommunizieren. Diese "Alzheimer Insektizide" unterbinden, dass die Sprache der Blumen von ihren Bestäubern weiterhin verstanden wird! Aber noch können wir Menschen gegensteuern!
Claudia Erbar & Peter Leins (Universität Heidelberg, Centre for Organismal Studies)
Erbar, C. & Leins, P. 2019: Ästhetische Ökonomie in der Gestalt der Pflanzen. – HDJBO 4: 39–90
Erbar, C. & Leins, P. 2023: Schöpferischer Kampf und Kooperation im Licht der Organismischen Evolution. – HDJBO 8: 29–73